
Centaurea
sect. Jacea

|
| Centaureasect. Jacea |
|
Chrpy ze sekce Jacea patří mezi nedostatečně probádané, tzv. kritické skupiny středoevropské flóry. Špatně se určují kvůli celkové podobnosti jednotlivých taxonů a nedostatku vhodných určovacích znaků. Navíc jsou zde zastoupeny dvě ploidní úrovně, jednotlivé taxony se hojně kříží a jsou také poměrně plastické ve vztahu k podmínkám prostředí, což obojí vše znesnadňuje určování. Komplikovaná taxonomická struktura pak s sebou (jak je v těchto případech obvyklé) nese také značný zmatek v nomenklatuře. Zároveň jsou chrpy rostliny nápadné a krásné a jsou tak velmi atraktivní skupinou pro studium.
Chrpami se zabývám od roku 2000, kdy jsem začal pracovat na diplomové práci zaměřené na morfologickou a karyologickou variabilitu skupiny tzv. chrp parukářek (Centaurea phrygia agg.) v Česku. Postupně se studium rozšířilo jak geograficky (zejména do Karpat), tak co se týče studovaných otázek (např. hybridizace a její vztah k plodii, morfologická plasticita, nomenklatura) i množství studovaných taxonů.
Hlavní témata, kterými se v současnosti zabývám, jsou následující:

Hlavním znakem pro určování chrp ze sekce Jacea je tvar přívěsků zákrovních listenů (hnědá nebo černá suchomázdřitá část nasedající nahoře na zelenou část listenu, viz obr.). Podle něj (ale nejen podle něj) lze středoevropské taxony rozdělit do několika dobře vyhraněných skupin: Centaurea jacea agg., Centaurea nigrescens agg., Centaurea nigra, Centarea macroptilon + Centaurea oxylepis, Centaurea phrygia agg. Tyto skupiny se dají poznat snadno, ale rozlišení taxonů uvnitř nich je obvykle obtížné.
Nejvíce se věnuji skupině "parukářek" (Centaurea phrygia agg.). Postupně se povedlo shromáždit 79 populačních vzorků (většinou 30 jedinců z populace) z Česka, Slovenska, Polska, Ukrajiny, Rumunska a Ruska, na kterých jsem měřil celkem 22 znaků. Výsledky byly zpracovány "klasickým" postupem: shlukové analýzy a PCA na úrovni populací --> definice skupin na jejich základě --> diskriminační analýzy na úrovni jedinců testující možnost odlišení takto definovaných skupin.
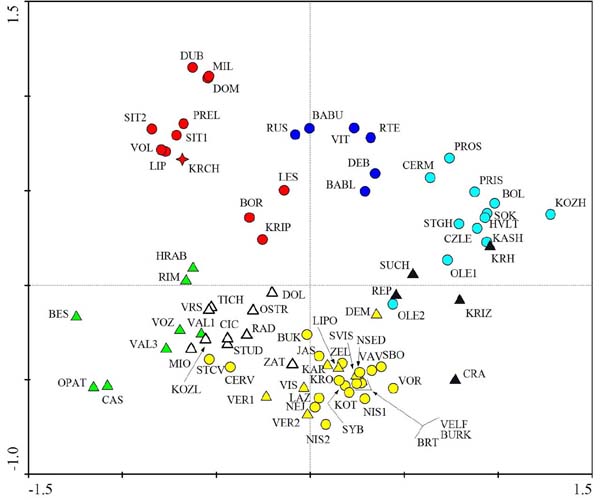 Celkem se povedlo rozlišit ve studovaném materiálu 5 taxonů, pro něž je vhodné použít kategorii druhu: Centaurea phrygia L. s. str., C. elatior (Gaud.) Hayek, C. carpatica Porc., C. stenolepis A. Kerner, C. oxylepis (Wimm. et Grab.) Hayek. V rámci C. phrygia L. existují dva cytotypy, které jsou dobře odlišené geograficky a do jisté míry i morfologicky a je možné rozlišovat je jako poddruhy subsp. phrygia a subsp. erdneri (Wagner) Koutecký. Dále roste na Slovensku (a dál na jihovýchod) C. indurata Janka, která ale nebyla v morfometrické analýze dostatečně zastoupena. Kromě "čistých" populací jednotlivých taxonů lze v přírodě najít také populace, které kombinují znaky dvou taxonů a jde zřejmě o jejich křížence.
Celkem se povedlo rozlišit ve studovaném materiálu 5 taxonů, pro něž je vhodné použít kategorii druhu: Centaurea phrygia L. s. str., C. elatior (Gaud.) Hayek, C. carpatica Porc., C. stenolepis A. Kerner, C. oxylepis (Wimm. et Grab.) Hayek. V rámci C. phrygia L. existují dva cytotypy, které jsou dobře odlišené geograficky a do jisté míry i morfologicky a je možné rozlišovat je jako poddruhy subsp. phrygia a subsp. erdneri (Wagner) Koutecký. Dále roste na Slovensku (a dál na jihovýchod) C. indurata Janka, která ale nebyla v morfometrické analýze dostatečně zastoupena. Kromě "čistých" populací jednotlivých taxonů lze v přírodě najít také populace, které kombinují znaky dvou taxonů a jde zřejmě o jejich křížence.
Do budoucna zbývá vyřešit ještě několik problémů ve střední Evropě, např. rozlišení C. oxylepis a C. macroptilon a možné odlišení dvou taxonů (panonského a karpatského) v rámci C. stenolepis. Téměř neznámá ale zůstává tato skupina na Balkáně, kde zbývá k objevení ještě mnoho. Stejně tak by bylo dobré získat nějaký materiál C. pseudophrygia z lesostepní zóny Ukrajiny a Ruska a srovnat ho se středoevropskými rostlinami označovanými také tímto jménem (pravděpodobně nejsou stejné).
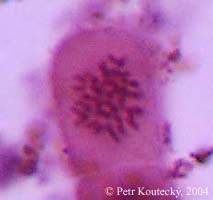
V rámci sekce Jacea se vyskytují dva chromozómové počty odpovídající dvěma ploidním úrovním: 2n = 22 (diploidní) a 2n = 44 (tetraploidní). Zatím se zdá, že jednotlivé taxony jsou v počtu chromozómů homogenní a chromozómový počet lze tak použít k rozlišení některých příbuzných (resp. morfologicky podobných) taxonů. Pro některé taxony, např. C. phrygia, C. nigra jsou sice udávány počty oba, ale je pravděpodobné, že jde o důsledek nevyřešených taxonomických problému a že v budoucnu bude vhodné rozlišit tyto "cytotypy" jako samostatné taxony. Příkladem je C. phrygia, u které se ukázalo, že cytotypy jsou odděleny geograficky i morfologicky a izolovány geneticky (právě odlišným počtem chromozómů) a je vhodné odlišovat je jako samostatné poddruhy, které jsou již v počtu chromozómů vnitřně homogenní.
Chromozómové počty jsou důležitým doplňkem pro morfometrickou analýzu a budou studovány i v budoucnu. Pro některé taxony jsou navíc neznámé nebo exitují v literatuře rozporné údaje, opět zejména v oblasti Balkánu.
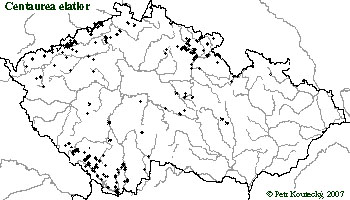
Geografické rozšíření i ekologické nároky jednotlivých morfologicky (příp. i karyologicky) definovaných "typů" jsou podstatnými faktory pro jejich taxonomické hodnocení. V rámci chrpy ze sekce Jacea se ukazuje, že morfologie a rozšíření a ekologie se dobře doplňují. Jednotlivé taxony mají středně velké až malé areály, které jsou dosti vyhraněné - areály jednotlivých taxonů v rámci jednotlivých druhových agregátů se téměř nepřekrývají.
Analýza rozšíření jednotlivých taxonů chrp je celkem obtížná. Jde o skupinu rostlin, které se špatně určují, a proto je v literatuře velké množství chybných údajů (vzniklých na základě chybného určení). Proto nelze literární údaje automaticky přebírat a všechny analýzy rozšíření mohou být založeny pouze na revizi herbářového materiálu nebo lokalit přímo v terénu. To je ale samozřejmě velmi časově náročné.
Pokud jde o ekologické nároky, většina taxonů roste ve střední Evropě nejčastěji na polopřirozených stanovištích jako jsou louky a pastviny, obvykle spíše na osluněných místech a ve středu gradientu vlhkosti (tj. ani vyslovená bažina ani step). Tyto taxony vstupují často i do lesních lemů nebo na ruderální stanoviště se spíše nízkou intenzitou narušování. Pouze několik taxonů roste v zásadně odlišných podmínkách. Nížinné taxony C. stenolepis a C. indurata rostou také na světlých místech v teplomilných doubravách (okraje, zarůstající paseky, staré lesní cesty) a na analogických stanovištích jako jsou křovité meze. Naopak C. carpatica je taxon vysloveně horský, který roste ve vysokostébelných společenstvech (vysokostébelné nivy kolem potoků a mezi porosty kleče, lavinové dráhy, apod.) nad horní hranicí lesa.

Taxonomická složitot chrp je jednou z příčin komplikované nomenklatorické situace. Jeden taxon je různými autory nazýván různě a naopak jedno jméno je různými autory různě vykládáno a přiřazováno k různým taxonům. V nastalém zmatku je obtížné se vyznat. Ve většině případů jediná cesta, jak zjistit, co dané jméno skutečně znamená, je revize původního popisu taxonu a zejména typového materiálu (obvykle stará, zaprášená a hmyzem poněkud sežraná položka). Práce je to někdy takřka detektivní a kromě prohledávání starých herbářů (obvykle někde v cizině, např. ve Vídni nebo Budapešti) znamená také luštění staré literatury, psané často v těžko srozumitelných jazycích (kromě latiny a ve střední Evropě všudypřítomné němčiny jsou některé důležité popisy taxonů chrp např. ukrajinsky, rumunsky nebo maďarsky).
V minulých letech jsem intenzívně studoval některá jména, která jsou často používána a jejichž skutečný obsah byl nejasný - mimo jiné Centaurea decipiens f. subjacea Beck (a další Beckova jména), C. melanocalathia Borbás, C. erdneri Wagner (a další jména J. Wagnera, zejména pro různé křížence).
Některé významné nomenklarotické otázky, na které jsem v průběhu studia narazil a kterými jsem se začal zabývat, zůstávají stále otevřené: typifikace Linného jména Centaurea phrygia, na které závisí nomenklatura poddruhů v rámci tohoto druhu a vůbec nomenklatura celé skupiny; správné jméno pro středoevropské rostliny nazývané buď Centaurea pseudophrygia C.A.Meyer nebo C. elatior (Gaud.) Hayek.; význam jmen Centaurea indurata Janka a C. nigriceps Dobrocz.; aj.

Vzhled chrp závisí podle mých zkušeností z terénu do značné míry na prostředí. Nejvýraznější vliv má silné zastínění a kosení. Zastíněné rostliny mají širší listy a jsou méně větvené než nezastíněné. Ještě výraznější rozdíly jsou mezi rostlinami první generace (vykvetlé před pokosením nebo na nekosených stanovištích) a rostlinami druhé generace (vyrostlé v létě nebo na podzim po pokosení). Rostliny druhé generace mají většinou méně větví, které jsou spíše vzpřímené a ne odstálé, dále mají většinou výrazně širší listy, ale zároveň delší internodia (jsou řídce olistěné), mívají řidší odění (nebývají šedivé) a určité rozdíly jsou i na úborech (viz obrázek, na kterém je právě taková "pokosená" rostlina). Určitý vliv na morfologii rostlin má i vlhkost a oslunění stanoviště (zejména na větvení rostlin, šířku listů a odění).
Jak je vidět, jsou prostředím ovlivněny zejména (i když ne výhradně) vegetativní znaky. Zároveň jsou v některých případech právě na základě těchto znaků rozlišovány některé taxony, např. poddruhy C. jacea nebo C. nigra.
V případě lépe známého případu C. jacea jsou ve střední Evropě tradičně rozlišovány dva poddruhy (podle některých autorů druhy): subsp. jacea a subsp. angustifolia (=C. pannonica). První z nich má být spíše chladnomilný a vázaný spíše na hor a podhůří, zatímco druhý má být spíše teplomilný, nížinný. Zároveň ve středních nadmořských výškách existují přechodné formy. Vzniká tak otázka, nejde-li spíše než o dva samostatné taxony pouze o projev morfologické plasticity taxonu jediného, široce rozšířeného.
Otázka morfologické plasticity by měla být jedním z hlavních témat studia sekce Jacea v blízké budoucnosti. Proto jsou na ni také vvpsána dvě témata bakalářských prací. Nabízejí se dva navzájem se doplňující metodické přístupy: srovnání morfologické a genetické variability (jde-li v případě C. jacea o dva taxony, měly by být odlišné nejen morfologicky, ale i geneticky) a přesazovací pokusy a studium populací v různých typech prostředí (jde-li o dva taxony, měly by být jejich znaky ±stálé i v různých prostředích).

Hybridizace je v sekci Jacea velmi častá. S trochou nadsázky lze říct, že v minulosti byli popsáni kříženci mezi skoro všemi druhy sekce ("každý s každým"). Kříženci jsou často plodní a kříží se zpětně s rodiči nebo mezi sebou nebo dokonce s některým třetím taxonem. Vznikají tak složité hybridní roje a dokonce dochází ke vzniku vícenásobných kříženců.
Na základě morfologických znaků je křížence obtížné určovat. Ve většině znaků leží "na půl cesty" mezi rodiči. Rodičovské taxony jsou ale navzájem dosti podobné a ještě víc to platí pro křížence. Nejnápadnější jsou kříženci mezi C. jacea jako jedním z rodičů a některým jiným taxonem jako druhým. Tito kříženci mají přívěsky zákrovních listenů nepravidelného tvaru s bočními třásněmi nepravidelně srostlými (viz obrázek) a vzhledem k hojnosti C. jacea jsou v populacích ostatních taxonů dosti častí.
Výskyt kříženců nebyl dřívějšími autory někdy brán dostatečně v úvahu (tj. nebyli rozpoznáni coby kříženci) a naopak někteří autoři intenzitu hybridizace přeceňovali a popisovali jako křížence nejrůznější netypické rostliny (ve skutečnosti nehybridní). To vede k tomu, že některé taxony uváděné tradičně jako nehybridní ve skutečnosti neexistují a jde o křížence (např. C. subjacea a C. phrygia subsp. melanocalathia a stejně tak neexistují některé udávané hybridní kombinace. To silně komlikuje zejména nomenklaturu celé skupiny (u každého jednotlivého jména je třeba vyzkoumat, jestli to není nějaký takový případ).
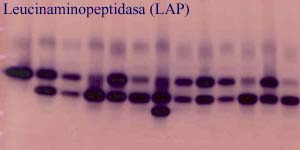 Kromě toho se zdá, že v některých případech dochází díky opakované hybridizaci k introgresi znaků jednoho taxonu do popu lací jiného. Výsledkem je, že v určitých územích nacházíme populace, které nelze přiřadit k žádnému z rozlišovaných taxonů a které v sobě kombinují znaky dvou taxonů (nejde ale o přímé křížence, protože se vyskytují samostatně, bez rodičovských taxonů, celá populace je tvořená pouze hybridními rostlinami). Například na celé severní Moravě jsou nejčastější chrpou rostliny, které morfologicky stojí mezi C. oxylepis a C. phrygia. Podobně to v části Podkrkonoší nebo v Železných horách platí pro populace přechodné mezi C. elatior a C. stenolepis a v Orlických horách se vzácně vyskytují rostliny ležící mezi C. elatior a C. phrygia. Potvrzení hybridního původu těchto rostlin je dalším úkolem do budoucna. Za prvé je třeba pomocí kontrolovaných pokusů ověřit, zda se uvedené druhy (předpokládané jako rodičovské) opravdu mohou křížit. Za druhé je potom nutné najít nějaký znak jiný než morfologický, který by dokázal, že morfologicky netypické populace jsou skutečně kříženci a ne jen extrémy variability jednoho z taxonů - v případě C. elatior - C. stenolepis byly testovány isoenzymy, ale zdá se, že se mezi uvedenými druhy moc neliší a že by bylo lepší hledat jiný genetický marker, založený přímo na analýze DNA.
Kromě toho se zdá, že v některých případech dochází díky opakované hybridizaci k introgresi znaků jednoho taxonu do popu lací jiného. Výsledkem je, že v určitých územích nacházíme populace, které nelze přiřadit k žádnému z rozlišovaných taxonů a které v sobě kombinují znaky dvou taxonů (nejde ale o přímé křížence, protože se vyskytují samostatně, bez rodičovských taxonů, celá populace je tvořená pouze hybridními rostlinami). Například na celé severní Moravě jsou nejčastější chrpou rostliny, které morfologicky stojí mezi C. oxylepis a C. phrygia. Podobně to v části Podkrkonoší nebo v Železných horách platí pro populace přechodné mezi C. elatior a C. stenolepis a v Orlických horách se vzácně vyskytují rostliny ležící mezi C. elatior a C. phrygia. Potvrzení hybridního původu těchto rostlin je dalším úkolem do budoucna. Za prvé je třeba pomocí kontrolovaných pokusů ověřit, zda se uvedené druhy (předpokládané jako rodičovské) opravdu mohou křížit. Za druhé je potom nutné najít nějaký znak jiný než morfologický, který by dokázal, že morfologicky netypické populace jsou skutečně kříženci a ne jen extrémy variability jednoho z taxonů - v případě C. elatior - C. stenolepis byly testovány isoenzymy, ale zdá se, že se mezi uvedenými druhy moc neliší a že by bylo lepší hledat jiný genetický marker, založený přímo na analýze DNA.
Chrpy ze sekce Jacea jsou polyploidní komplex se dvěma úrovněmi ploidie - diploidní a tetraploidní. To ovlivňuje některé jejich vlastnosti, mimo jiné i hybridizaci. Taxony chrp o stejné ploidii (jedno jaké) se kříží velmi dobře a kříženci jsou obvykle plodní, zatímco kříženci taxonů různé ploidie jsou vzácní a téměř sterilní. Různá ploidní úroveň tak přestavuje významnou reprodukční bariéru, zřejmě jednu z mála, ne-li jedinou, které v této skupině rostlin existují (samozřejmě kromě geografické izolace).
Kříženci mezi diploidy a tetraploid vzniklí normální cestou by měli být triploidní. Takové rostliny se ale u chrp v přírodě téměř nevyskytují a i v cílených hybridizačních experimentech jsou dosti vzácné. Kromě nich se ale vzácně objevují (v přírodě i v pokusech) tetraploidní kříženci, kteří vznikají za účasti neredukovaných gamet diploidního rodiče. Narozdíl od triploidů bývají plodní a mohou se účastnit dalšího křížení (na tetraploidní úrovni).
Výskyt neredukovaných gamet je velmi významným faktorem pro fungivání polyploidních komplexů. Na základě výsledků teoretických modelů se zdá, že může jít o klíčový faktor umožňující vznik a dlouhodobé přežívání tetraploidů. Empirických dat potvrzujících tyto modely je ale velmi málo. Chrpy ze sekce Jacea se zdají být pro tyto studie ideální modelovou rostlinou (víme, že tam k studovanému jevu dochází, a navíc jsou v mnohých ohledech zcela standardní - téměř nemají vegetativní rozmnožování, semena vznikají normálním pohlavním rozmnožováním s téměř striktní alogamií, jsou opylovány hmyzem a tak lze uvažovat o přenosu gamet zejména v rámci jedné prostorově omezené populace, apod.). Proto se na studium hybridizačních procesů u chrp v budoucnu jistě zaměřím, i téma pro šikovného a pečlivého studenta (zejména pokusy vyžadují pečlivost a trpělivost) by se našlo.